
本学 橋本敬一郎名誉教授らの研究成果が米国科学アカデミー紀要「Proceedings of the National Academy of Sciences of the United States of America」に掲載されました
主要組織適合遺伝子複合体(MHC)分子の出現は
クラスI・IIのどちらが先か─。
免疫学者の間で数十年来の謎
クラス分岐の進化過程に重要な3つ目のカテゴリーを発見
クラスI・IIのどちらが先か─。
免疫学者の間で数十年来の謎
クラス分岐の進化過程に重要な3つ目のカテゴリーを発見
本学 岡村和彦・元医療科学部准教授、ヨハネス=ダイクストラ・総合医科学研究所准教授、塚本健太郎・研究支援推進本部講師および橋本敬一郎・名誉教授・元総合医科学研究所医高分子学研究部門教授の研究グループらは、これまでクラスIかクラスIIに分類されてきた主要組織適合遺伝子複合体(MHC)分子のカテゴリーに、古くから存在する3つ目のカテゴリー「MHC-W分子群」を発見しました。
私達の健康に重要な免疫機構において、Tリンパ球に抗原を提示する重要な機能を有する主要組織適合遺伝子複合体(MHC)分子は、これまでカテゴリーとしてクラスIかクラスIIに分類されてきました。MHCクラスI分子※1は、ウイルスや癌細胞などを排除する細胞傷害性T細胞が関わる機構に重要であり、MHCクラスII分子※1は、ヘルパーT細胞が関わる様々な抗原に対応する為の効率的な抗体産生機構などに重要です。この様に、MHC分子のクラス分岐は、私達の免疫機構の基礎を担っていますが、その成立に関しては謎でした。MHC-W分子群は、クラスIに近い配列を有しますが、クラスIIと同様に似たサイズのアルファ(α)鎖とベータ(β)鎖を有しています。この発見は、クラスIとクラスIIのどちらが先に現れたかという分子進化の問題において、クラスII型のα鎖・β鎖の構成を持つ分子が最初に出現した結論を導き、数十年に渡り免疫学者にとって謎であった問題を解明することに至りました。
本研究成果は、米国科学アカデミー紀要「Proceedings of the National Academy of Sciences of the United States of America(PNAS)」(12月21日号)の掲載に先駆け、オンライン版が12月14日5時00分(JST)に公開されました。
研究成果のポイント
- MHCクラスIとMHCクラスIIの2種以外に新しいカテゴリーとしてMHC-Wを発見
- MHC-Wは、MHC-IIとクラスII型のドメイン構成を共有している一方、MHC-Iとはドメイン間モチーフ配列を共有し、また系統樹解析で近縁であることを解明
- 得られた結果に基づき、MHC-WからMHC-Iが生じ、MHCクラス分岐※2の謎に対してクラスII型が先に出現した分子進化を提唱
背景
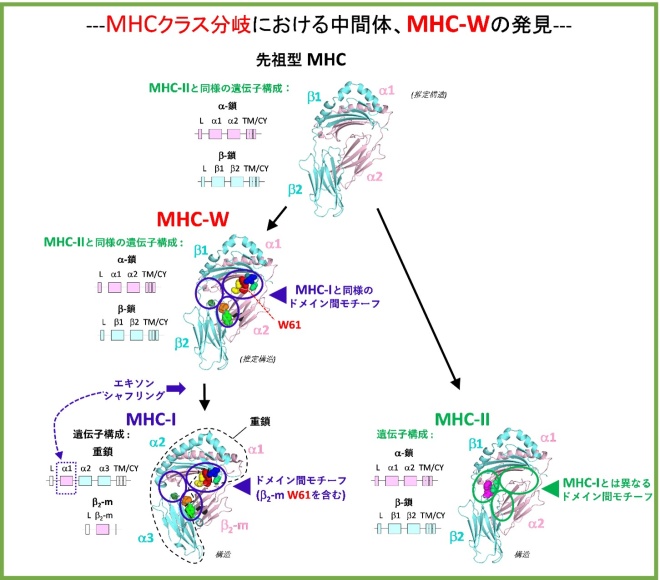
どちらが先に出現したのか、MHCクラスIかMHCクラスIIか?──免疫学者の間ではこれらの2つのクラスのうち、分子進化上、どちらが先に現れたかが数十年に渡り議論されてきました。この度、橋本研究グループは、ヨーロッパの研究者らの協力も得て、米国科学アカデミー紀要に発表される新しい論文において、この疑問に対する答を得ることができました(文献1)。著者らは、MHC分子の古くから存在するカテゴリーであるMHC-Wを発見しましたが、この分子は、クラスI分子が元のクラスII分子からいかにして進化したかを説明しうる、これまで不明であった中間的分子であることが判りました。今回、米国科学アカデミー紀要のゲスト編集者として本論文を担当したスタンフォード大学のピーター・パーハム教授は、次の様に述べています。「MHCクラスIIがMHCクラスIに先行して進化したこと、そしてそのMHCクラスIはMHCクラスIIから進化したことを論証することによって、この画期的な研究は、免疫遺伝学における重要で非常に難解なパズルを解明しました。」MHCクラスI分子とクラスII分子は、互いに密接に関係しています。どちらのタイプも4つの細胞外ドメインを有しており、そのうち細胞膜より遠い所に位置する2つのドメインは、Tリンパ球へ提示されるペプチド抗原を結合する溝構造を形成しています。しかしながら、クラスI分子とクラスII分子では、ドメインの連結様式が異なります。クラスII分子は、2つの細胞外ドメイン(II-α1とII-α2)を有するα鎖と、2つの細胞外ドメイン(II-β1とII-β2)を有するβ鎖の、似たサイズの2種類の細胞膜結合型の鎖から成っています。一方、クラスI分子は、3つの細胞外ドメイン(I-α1、I-α2、I-α3)を有する大きな細胞膜結合型のα鎖と、ただ一つのドメインから成る遊離のβ2-ミクログロブリン(β2-m)から形成されています(図1)。構造的に比較すると、次のクラスIおよびクラスII分子のドメインが、それぞれ対応する関係になっています。即ち、II-α1とI-α1、II-α2とβ2-m、II-β1とI-α2、そしてII-β2とI-α3の組み合せになります(図1)。ドメイン相互の類似性および分子進化における簡潔性を考慮することにより、ジム・カウフマン教授と共同研究者は既に1984年に、分子進化上MHC分子はホモダイマーとして出現し、遺伝子重複および分化を経てクラスII型のヘテロダイマーとなり、それが現在のクラスIおよびクラスIIの起源となったであろうことを提唱しています(文献2、図1)。しかしながら、彼らの結論は論争を引き起こし(文献3)、今日に至るまで、どちらのクラスが先に出現したのかは、解明されてきませんでした。重要なこととして、MHCクラスの分子進化の解明に光を当て得る、クラスIおよびクラスII両者の性質を有する原始的なMHCグループが見い出されてこなかったことが挙げられます。
研究手法・研究成果
ジム・カウフマン教授が1984年に得ることができたものに比べ、橋本研究グループは、遥かに多様な動物種から多くのMHC配列を得て、より信頼できる系統樹解析を実施することができる様になり、カウフマン教授のモデルを支持する結果を得ることができました。最も重要なことには、サメ類等の軟骨魚類、条鰭類や肉鰭類の硬骨魚類、両生類のサラマンダー等を含む原始的な有顎脊椎動物より、新しいカテゴリーに属するMHC分子としてMHC-Wを発見し、これはクラスIに近い配列を有し、同様なサイズの細胞膜結合型α鎖およびβ鎖から構成されていることを明らかにしました(図1)。この発見は、クラスII型の細胞膜結合型α鎖およびβ鎖を有するMHC分子が分子進化上最初に出現したことを結論しており、カウフマン教授のモデルに最終的な証拠を与えることになりました。詳細な配列解析からは、ドメイン間の結合配列モチーフが、MHC-IとMHC-WをMHC-IIから区別する重要な特徴となっています。MHC-Wは、MHC-Iの重鎖がβ2-mとの結合において重要な配列モチーフを有しており、その配列中にはβ2-mとMHC-Wのα2の両者で保存されていてMHC-Wがその名を得た重要なトリプトファン(アミノ酸の一文字表記でW)が含まれています(図1)。このことは、遺伝子構成を変化させるエキソン・シャフリング※3機構が先祖としてのMHC-WからMHC-Iの重鎖とβ2-mを生じた以前に、既にMHC-Wの段階でMHC-I重鎖とβ2-mとの結合機構が進化していたことを示唆しています(図1)。
元ケンブリッジ大学教授で現在エジンバラ大学教授のジム・カウフマン教授は、本研究について、次の様なコメントを寄せています。「米国科学アカデミー紀要に掲載予定の岡村らの研究は、長年に渡り哺乳類以外の脊椎動物から集められた様々な種類のデータにより強力に支持される全く新しい予想外のMHC遺伝子ファミリーを明らかにした大作です。橋本研究グループによって提唱されたシナリオは、MHC遺伝子の初期進化を理解する上で、数十年で初めての進歩です。」
MHC-Wは、MHC分子の分子進化において、クラスII型の分子構成が最初に出現した証拠を提供する以外に、将来、MHC研究領域における長年の謎の一つであるβ2-m機能の仕組みを解明するのに役立つ可能性が考えられます。
参考文献
- Okamura K, Dijkstra JM, Tsukamoto K, Grimholt U, Wiegertjes GF, Kondow A, Yamaguchi H, Hashimoto K. Discovery of an ancient MHC category with both class I and class II features. PNAS https://doi.org/10.1073/pnas.2108104118
- Kaufman JF, Auffray C, Korman AJ, Shackelford DA, Strominger J (1984) The class II molecules of the human and murine major histocompatibility complex. Cell 36(1):1-13.
- Flajnik MF, Canel C, Kramer J, Kasahara M (1991) Which came first, MHC class I or class II? Immunogenetics. 1991;33(5-6):295-300.
用語解説
※1 MHCクラスI分子、MHCクラスII分子
主要組織適合遺伝子複合体(major histocompatibility complex, MHC)分子は、歴史的には、移植反応の時の組織適合性に関わる分子として見つかりました。MHC分子は、免疫反応においてペプチド抗原をT細胞へと提示する重要な機能を有し、MHCクラスI分子とMHCクラスII分子に大別されます。MHCクラスI分子は、ヒトでは、HLA-A、HLA-B、HLA-Cがあり、ほとんどの有核細胞に発現しています。CD8陽性のキラーT細胞のT細胞レセプターへ主に内因性ペプチド抗原を提示し、ウイルスや癌細胞の排除に関与します。またMHCクラスII分子は、ヒトではHLA-DP、HLA-DQ、HLA-DRがあり、B細胞、樹状細胞、マクロファージなどに発現しています。CD4陽性のヘルパーT細胞のT細胞レセプターへ主に外来性ペプチド抗原を提示し、強力な抗体産生の補助等に重要な役割を担っています。※2 MHCクラス分岐
MHCクラスI分子とMHCクラスII分子は、アミノ酸配列や立体構造が大きく類似しています。しかしながら、ドメイン連結構成に大きな違いがあります。それぞれ4つから成る細胞外ドメインを見ると、MHCクラスI分子は、3ドメインが連結していて、あと遊離型のドメイン(β2-ミクログロブリン、β2-m)が1つ結合してダイマーを作り計4ドメインとなります。一方、MHCクラスII分子は、2ドメインが連結して1つの鎖を形成しており、αとβの2種の鎖がダイマーを作り、計4ドメインとなります。分子進化上MHCクラスI分子とMHCクラスII分子のどちらが先に現れたかについては、1984年以降、クラスII先行説、クラスI先行説が提唱されてきましたが、分子進化の方向性の示唆に重要であろう中間体などが全く得られていなかったため、明確な答は出ていませんでした。※3 エキソン・シャフリング
成熟mRNAに残る塩基配列部分であるエキソンが単位となって、色々な組み合わせの変化を起こして新しい遺伝子を生じるであろう分子機構。文献情報
論文タイトル
Discovery of an ancient MHC category with both class I and class II features著者
岡村 和彦1*, ヨハネス M.ダイクストラ2*, 塚本 健太郎3*, Unni Grimholt4, Geert F. Wiegertjes5, 近藤 晶子6, 山口 央輝7, 橋本 敬一郎8§所属
1 元藤田医科大学医療科学部臨床工学科2 藤田医科大学総合医科学研究所
3 藤田医科大学研究支援推進本部共同利用研究設備サポートセンター・タンパク質遺伝子解析室
4 Fish Health Research Group, Norwegian Veterinary Institute、ノルウェー
5 Aquaculture and Fisheries Group, Department of Animal Sciences, Wageningen University & Research、オランダ
6 帝京大学先端総合研究機構
7 四日市看護医療大学看護医療学部臨床検査学科
8 藤田医科大学名誉教授・元総合医科学研究所医高分子学研究部門
* 共同第一著者 § 責任著者
DOI
10.1073/pnas.2108104118問い合わせ先
総合医科学研究所
准教授 ヨハネス M. (ハンス)ダイクストラ
e-mail:Dijkstra@fujita-hu.ac.jp




